

BY: K. Sai Manogna (MSIWM014)
At inflammatory sites and secondary lymphoid glands, different subsets of lymphocytes exhibit directed extravasation. Therefore, lymphocyte recirculation is closely monitored to ensure that sufficient populations of B and T cells are recruited into various tissues. Extravasation of lymphocytes involves interactions between a variety of cell-adhesion molecules, as with neutrophils. The overall process is similar to what occurs during the extravasation of neutrophils and involves the same four stages of touch and rolling, activation, arrest and adhesion and, eventually, transendothelial migration.
Sites of Lymphocyte Extravasation:
1. Some regions of vascular endothelium consist of specialised cells with a plump, cuboidal (‘high’) form in the postcapillary venules of different lymphoid organs; such regions are referred to as high-endothelial venules or HEVs.
2. In appearance, their cells contrast strongly with the flattened endothelial cells that line the rest of the capillaries. Each of the secondary lymphoid organs comprises HEVs, except the spleen.
3. There are about 1.4 × 104 lymphocytes extravasate into a single lymph node every second through HEVs.
4. Cytokines developed in response to antigen capture influence the production and maintenance of HEVs in lymphoid organs.
5. In order to prevent the antigen from entering the node, the role of lymphocyte antigenic activation in the preservation of HEVs has been demonstrated by surgical blocking of afferent lymphocyte vasculature of the node.
6. The HEVs demonstrate impaired function within a short period and gradually return to a more flattened morphology.

7. High-endothelial venules express several cell-adhesion molecules. HEVs, like other vascular endothelial cells, express CAMs from the selectin (E- and P-selectin) family, the mucin-like (GlyCAM-1 and CD34) family, and the superfamily of immunoglobulins (ICAM-1, ICAM-2, ICAM-3, VCAM-1, and MAdCAM-1).
8. In a tissue-specific way, some of these adhesion molecules are distributed. These tissue-specific adhesion molecules have been named vascular addressins (VAs) because they help to guide the extravasation to specific lymphoid organs of various populations of recirculating lymphocytes.
Receptor Profiles and Signals Guided by Lymphocyte Homing:
1. Related to neutrophil extravasation, the general lymphocyte extravasation mechanism is similar.
2. The fact that different subsets of lymphocytes migrate differently into different tissues is a significant aspect that separates the two processes. This method is known as trafficking or homing.
3. The numerous lymphocyte subset trafficking patterns are regulated by unique combinations of adhesion molecules and chemokines; homing receptors are called receptors that guide the circulation of different lymphocyte populations to specific lymphoid and inflammatory tissues.
Researchers have established several lymphocytes and endothelial cell adhesion molecules that are involved in lymphocyte interactions with HEVs and endothelium at tertiary sites or sites of inflammation.
Recirculating Naive Lymphocytes into Secondary Lymphoid Tissue:
Until it has been triggered to become an effector cell, a naive lymphocyte is not able to mount an immune response.
1. In specialised microenvironments within secondary lymphoid tissue (e.g., peripheral lymph nodes, Peyer patches, tonsils, and spleen), activation of a naive cell occurs.
2. Dendritic cells catch antigen inside these microenvironments and present it to the naive lymphocyte, resulting in its activation.
3. Naive cells do not display a preference for a specific form of secondary lymphoid tissue but instead circulate indiscriminately across the body to secondary lymphoid tissue through recognising HEV adhesion molecules.
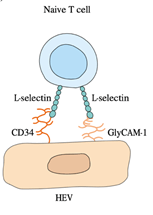
4. The initial attachment to HEVs of naive lymphocytes is usually mediated by the binding of the L-selectin homing receptor to HEV adhesion molecules such as GlyCAM-1 and CD34.
5. The naive cell trafficking pattern is designed to keep these cells continuously recirculating across secondary lymphoid tissue, the primary purpose of which is to trap antigen transmitted by blood or tissue.
6. They are activated and enlarged into lymphoblasts until naive lymphocytes encounter antigen trapped in secondary lymphoid tissue. Activation takes approximately 48 h, and the blast cells are retained in the paracortical area of secondary lymphoid tissue during this time.
7. The antigen-specific lymphocytes cannot be identified in the circulation during this process, called the shutdown phase.
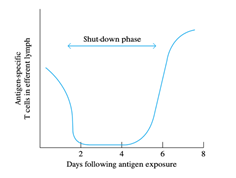
8. During the shutdown point, rapid proliferation and differentiation of naive cells occur. Then the effector and memory cells that this process produces leave the lymphoid tissue and begin to recirculate.
Lymphocytes of Effector and Memory follow distinct patterns of trafficking:
1. Effector and memory lymphocyte trafficking patterns vary from those of naive lymphocytes.
2. By recognising inflamed vascular endothelium and chemoattractant molecules produced during the inflammatory response, effector cells appear to be home to regions of infection.
3. On the other hand, memory lymphocytes selectively house the type of tissue in which antigen was first encountered.
4. This presumably ensures that a specific memory cell returns to the tissue where the antigen it recognises is most likely to re-encounter a subsequent threat.
5. Memory cells and effector cells express increased levels of specific molecules of cell adhesion, such as LFA-1, which interact with ligands present in additional tertiary lymphoid tissue (such as skin and mucosal epithelial) and at inflammation sites, allowing these sites to be accessed by effector and memory cells.
6. Naive cells lack the corresponding molecules of cell-adhesion and do not house these sites.
7. A variety of adhesion molecules, including E- and P-selectin and the Ig-superfamily molecules VCAM-1 and ICAM-1, are expressed in inflamed endothelium and bind to receptors expressed at high levels in the memory and effector cells.
8. Subsets of the memory and effector populations display tissue-selective homing activity, unlike naive lymphocytes.
9. Such tissue specificity is imparted by multiple combinations of adhesion molecules rather than by a single adhesion receptor.
10. A mucosal homing subset of memory/effector cells, for example, has high levels of LPAM-1 (4 7) and LFA-1 (Lb2) integrins that bind to MAdCAM and various ICAMs on venules of intestinal lamina propria.

11. However, since they have low levels of L-selectin that would promote their entry into secondary lymphoid tissue, these cells prevent direction to secondary lymphoid tissues.
12. Preferential homing to the skin is shown by the second group of memory/effector cells. Low levels of L-selectin are also expressed in this subset, however high levels of cutaneous lymphocyte antigen (CLA) and LFA-1, which bind to E-selectin and ICAMs on skin dermal venules, are seen.
13. While effector cells and memory cells that express decreased L-selectin levels do not appear to reach peripheral lymph nodes via HEVs, they may enter peripheral lymph nodes via afferent lymph vessels.
Adhesion-Molecule Interactions Play Extravasation Vital Roles:
1. A multi-stage mechanism involving a cascade of adhesion-molecule interactions is the extravasation of lymphocytes into secondary lymphoid tissue or regions of inflammation, similar to those involved in bloodstream neutrophil emigration.
2. This shows the usual interactions in the extravasation of naive T cells into lymph nodes through HEVs.
Mechanism:
1. In the first stage, a selectin-carbohydrate interaction similar to that seen with neutrophil adhesion.
2. L-selectin, which acts as a homing receptor which directs the lymphocytes to specific tissues expressing a corresponding mucin-like vascular addressin such as CD34 or GlyCAM-1, initially binds naive lymphocytes to HEVs.
3. The rolling of lymphocytes is less pronounced than neutrophil rolling.
4. Although the initial interaction of selectin-carbohydrate is minimal, the slow rate of blood flow in postcapillary venules, especially in regions of HEVs, reduces the possibility that the tethered lymphocyte can dislodge the sheer force of the flowing blood.
5. In the second stage, chemokines that are either localised on the endothelial surface or secreted locally mediate an integrin-activating stimulus.
6. To maintain these soluble chemoattractant variables on the HEVs, the thick glycocalyx covering of the HEVs can work.
7. If, as some have indicated, HEVs secrete lymphocyte-specific chemoattractants, it will clarify why, while they express L-selectin, neutrophils do not extravasate into lymph nodes at the HEVs.
8. As happens in neutrophil extravasation, chemokine binding to G-protein – coupled receptors on the lymphocyte contributes to the activation of integrin molecules on the membrane.
9. The integrin molecules interact with the adhesion molecules of the Ig superfamily (e.g., ICAM-1) once activated, so that the lymphocyte adheres tightly to the endothelium.
10. In the final stage, molecular mechanisms involved in the transendothelial migration are poorly understood.

