display MHC complexes in association with these peptide fragments of protein antigens on its surface to lymphocytes, and
provide signals that stimulate the proliferation and differentiation of the lymphocytes (co-stimulatory signal) are known as the antigen-presenting cells (APCs).
APCs are conventionally referred to as those cells which display antigens on their surface to the T lymphocytes. A dendritic cell is the major type of APC which is involved in initiating the T cell responses.
Macrophages and B cells also present antigens to the T lymphocytes but in different types of immune responses. The follicular dendritic cell, a specialized cell type, displays antigens to B lymphocytes during particular phases of humoral immune responses. APCs thus link responses of the innate immune system to responses of the adaptive immune system, and therefore they may be considered components of both systems.
Types of APCs:
Dendritic Cells
Dendritic cells form one of the most important APCs for activating naive T cells. These cells constitutively express a high level of class II MHC molecules and deliver a co-stimulatory activity and thus play major roles in innate responses to infections and link innate and adaptive immune responses.
Antigen-Presenting Cells for Effector T -Lymphocytes
In addition to dendritic cells; macrophages and B lymphocytes perform important antigen-presenting functions in CD4+ helper T cell-mediated immune responses.
Macrophages present antigen to helper T lymphocytes at the sites of infection, which leads to helper T cell activation and production of molecules that further activate the macrophages. These macrophages must be activated by phagocytosis of particulate antigens before expressing class II MHC molecules or the co-stimulatory B7 membrane molecule.
B cells present antigens to helper T cells in lymph nodes and spleen, a key step in the cooperation of helper T cells with B cells in humoral immune responses to protein antigens. These B cells constitutively express class II MHC molecules but must be activated before expressing the co-stimulatory B7 molecule.
Note: Cytotoxic T lymphocytes (CTLs) are effector CD8+ T cells that can recognize antigens on any nucleated cell and become activated to kill the cell. Thus, all nucleated cells are potentially APCs for CTLs.
Follicular Dendritic Cells:
Follicular dendritic cells (FDCs) are cells with membranous projections found intermingled in specialized collections of activated B cells, called germinal centers, in the lymphoid follicles of the lymph nodes, spleen, and mucosal lymphoid tissues.
Article by- SAMPRATI PAREKH (MSIWM049)
References:
Cellular and Molecular Immunology by Abul K. Abbas – 7thEdition
Each mammalian species studied to date has a closely linked gene cluster. The major histocompatibility complex (MHC) plays a role in intercellular recognition and self-nonself-discrimination. The MHC is involved in the production of immune responses that are both humoral and cell-mediated. While antibodies may respond on their own with antigens, most T cells only recognize antigens when combined with an MHC molecule. Also, since MHC molecules act as antigen-presenting structures, the unique collection of MHC molecules expressed by an individual influences the antigen repertoire to which the TH and TC cells of that individual may respond. For this reason, an individual’s response to antigens from infectious organisms is partly determined by the MHC and has therefore been involved in disease susceptibility and autoimmunity production.
General Organization and Inheritance of the MHC:
1. The theory that foreign tissue rejection is the result of an immune reaction to cell-surface molecules, now called antigens of histocompatibility, arose from Peter Gorer ‘s work in the mid-1930s.
2. To recognize blood-group antigens, Gorer used inbred strains of mice. He identified four groups of genes, designated I through IV, that encoded blood-cell antigens in the course of these studies.
3. Work carried out by Gorer and George Snell in the 1940s and 1950s established that gene-encoded antigens in the group designated II were involved in the rejection of transplanted tumors and other tissue. Snell called these genes “histocompatibility genes”; their current designation was about Gorer ‘s group II blood-group antigens as histocompatibility-2 (H-2) genes.
4. Although Gorer died before his contributions were fully recognized, Snell was awarded the 1980 Nobel Prize for this work.
A series of genes arranged inside a long, continuous stretch of DNA on chromosome 6 in humans and chromosome 17 in mice is the main histocompatibility complex. The MHC is referred to in humans as the HLA complex and in mice as the H-2 complex. Although the arrangement of genes is quite different, the MHC genes are grouped into regions encoding three groups of molecules in both cases.
Class I MHC Genes: Glycoproteins expressed on the surface of almost all nucleated cells are encoded by class I MHC genes; the critical feature of class I gene products is the presentation to TC cells of peptide antigens.
Class II MHC Genes: Glycoproteins expressed predominantly in antigen-presenting cells (macrophages, dendritic cells, and B cells) are encoded by class II MHC genes, where they present processed antigenic peptides to TH cells.
Class III MHC Genes: In addition to other products, Class III MHC genes encode different secreted proteins which have immune functions, including components of the complement system and molecules that are involved in inflammation.
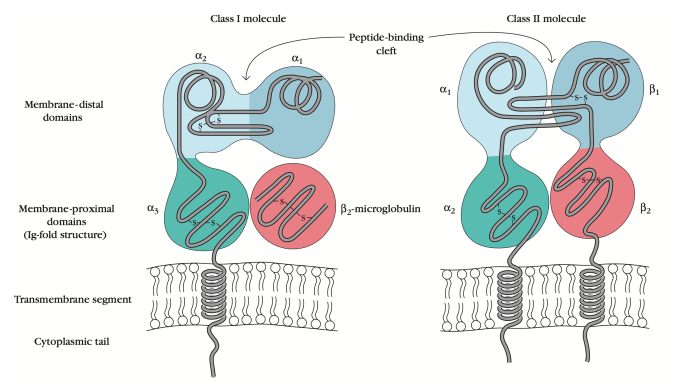
Both MHC molecules of class I and class II are membrane-bound glycoproteins closely related in both structure and function. These molecules have been isolated and purified, and x-ray crystallography has determined their extracellular domains’ three-dimensional structures. Both membrane glycoprotein types act as highly specialized antigen-presenting molecules that form extremely stable antigenic peptide complexes, displaying them on the cell surface for T cell recognition. MHC molecules of class III, on the other hand, are a group of unrelated proteins which do not share structural similarities and common function with molecules of class I and II.
Class I MHC Molecules:
1. A 45-kilodalton (kDa) alpha chain-linked noncovalently with a 12-kDa beta2-microglobulin molecule is found in Class I MHC molecules.
2. The alpha chain is a transmembrane glycoprotein encoded within the A, B, and C regions of the human HLA complex by polymorphic genes and within the K and D/L regions of H-2 complex of the mouse.
3. Beta2-Microglobulin is a protein encoded on a distinct chromosome by a strongly conserved gene.
4. For the expression of class-I molecules on cell membranes, the alpha chain association with beta2-microglobulin is required.
5. With its hydrophobic transmembrane part and hydrophilic cytoplasmic tail, the alpha chain is anchored in the plasma membrane.
Structure of Class I MHC:
1. Structural analyses have shown that the class I MHC alpha chain molecules are divided into three external domains (alpha1, alpha2, and alpha3. Each contains approximately 90 amino acids; a transmembrane domain-containing approximately 25 hydrophobic amino acids, followed by a short stretch of charged amino acids; and a 30 amino acid cytoplasmic anchor segment.
2. The beta2-microglobulin is similar in size and organization to the alpha3 domain; it does not contain a transmembrane region and is noncovalently bound to the class I glycoprotein.
3. Sequence data shows homology in immunoglobulins between the alpha3, beta2-microglobulin, and constant-region domains.
4. The enzyme papain cleaves the alpha chain just 13 residues proximal to its transmembrane domain, releasing the extracellular portion of the molecule, consisting of a1, a2, a3 beta2-microglobulin.
5. Two pairs of interacting domains were revealed by purification and crystallization of the extracellular portion: a membrane-distal pair composed of the alpha-1 and alpha-2 domains and a membrane-proximal pair composed of the alpha-3 domain and beta2-microglobulin.
6. The alpha-1 and alpha-2 domains combine to form a platform of eight antiparallel beta-strands spanned by two long alpha-helical regions.
7. With long alpha-helices as sides, and beta-strands of the beta-sheet as the bottom, the structure forms a deep groove, or cleft, about 25Å ×10Å ×11Å.
8. On the top surface of class I MHC molecule, this peptide-binding cleft is located, and it is wide enough to bind a peptide of 8-10 amino acids.
9. The discovery of tiny peptides in the cleft that had co-crystallized with the protein was the big surprise in the x-ray crystallographic study of class I molecules. These peptides are processed antigen and self-peptides bound to the alpha-1 and alpha-2 domains in this deep groove.
10. The a3 domain and beta2-microglobulin are organized into two beta-pleated sheets, each formed by antiparallel beta-strands of amino acids. As defined, this structure, known as the immunoglobulin fold, is characteristic of immunoglobulin domains.
11. Class I MHC molecules and beta2-microglobulin are known as immunoglobulin members due to this structural similarity.
12. Among class I MHC molecules, the alpha-3 domain appears highly conserved and contains a sequence interacting with the CD8 membrane molecule present on TC cells.
13. Beta2-Microglobulin deeply interacts with the alpha-3 domain and also interacts with the a1 and a2 domain amino acids.
For the Class I molecule to achieve its fully folded conformation, the interaction of beta2-microglobulin and a peptide with a class I alpha chain is necessary. The assembly of class I molecules is assumed to occur by the initial interaction of beta2-microglobulin with the folding class I alpha chain. The binding of a suitable peptide to form the native trimeric class I structure consisting of the class I alpha chain, beta2-microglobulin, and a peptide stabilizes this metastable “empty” dimer. Ultimately, this full molecular complex is transferred to the cell surface.
Class II MHC Molecules:
1. There are two distinct polypeptide chains in Class II MHC molecules, a 33-kDa alpha chain and a 28-kDa beta chain connected by noncovalent interactions.
2. Class II MHC molecules, including class I alpha chains, are membrane-bound glycoproteins containing external domains, a segment of the transmembrane, and a segment of the cytoplasmic anchor.
3. In a class II molecule, each chain comprises two external domains: the domains alpha-1 and alpha-2 in one chain and the domains beta-1 and beta-2 in the other.
4. Like the membrane-proximal alpha-3 / beta-2-microglobulin domains of class I MHC molecules, the membrane-proximal alpha-2, and beta-2 domains bear sequence resemblance immunoglobulin-fold structure; thus, class II MHC molecules are also known as super-family immunoglobulins.
5. The membrane-distal part of the class II molecule consists of the domain’s alpha-1 and beta-1 and forms the antigen-binding cleft for the antigen being processed.
6. The X-ray crystallographic analysis indicates that class II and class I molecules are identical, which is remarkably obvious when the molecules are superimposed.
7. Like that in class I molecules, the peptide-binding cleft of HLA-DR1 is composed of a floor of eight antiparallel beta-strands and sides of antiparallel alpha-helices.
8. The class II molecule lacks conserved residues that bind short peptides to the terminal residues and form an open pocket instead; class I offers more of a socket, class II, an open-ended groove.